Pombalia attenuata (Humb. & Bonpl. ex Willd.) Paula-Souza
Common names:
Western Green Violet
Synonyms:
Pombalia attenuata (Humb. & Bonpl. ex Willd.) Paula-Souza, in Paula-Souza and H.E.Ballard, Phytotaxa 183(1): 6. 2014; Hybanthus attenuatus (Humb. & Bonpl.) Schulze-Menz, Notizbl. Bot. Gart. Berlin-Dahlem 12: 114. 1934; Ionidium attenuatum Humb. & Bonpl., in Roem. & Schult., Syst. veg. 5: 402. 1819. TYPE: Columbia. Angostura, [no date], Humboldt & Bonpland 1643 (holotype: B-W!; isotype: P00604176!).
Hybanthus elatus (Turcz.) C.V.Morton, Contr. U. S. Natl. Herb. 29: 80. 1944; Ionidium elatum Turcz., Bull. Soc. Imp. Naturalistes Moscou 36(1): 556. 1863
Mercurialis glabra M.E.Jones, Contr. West. Bot. 18: preprint 68. 1933
Calceolaria riparia (Kunth) Kuntze var. houstonii (DC.) Dowell, Bull. Torrey Bot. Club 33: 554. 1906.; Ionidium parietariifolium DC. var. houstonii DC., Prodr. 1: 308. 1824
Hybanthus parietariifolius (DC.) Loes., Bull. Herb. Boissier, Sér. II, 3: 214. 1903; Solea parietarifolia (DC.) Spreng., Syst. veg. 1: 803-804. 1824; Ionidium parietariifolium DC., Prodr. 1: 308. 1824
Calceolaria riparia (Kunth) Kuntze, Revis. gen. pl. 1: 41. 1891; Solea riparia (Kunth) Spreng., Syst. veg. 1: 803. 1825 ["1824"]; Ionidium riparium Kunth, in Humb., Bonpl. & Kunth, Nov. gen. sp. 5(23)[quarto]: 378. 1823 [nomen superfluum]
Calceolaria botteri (Turcz.) Kuntze, Revis. gen. pl. 1: 41. 1891; Ionidium botteri Turcz., Bull. Soc. Imp. Naturalistes Moscou 36(1): 556. 1863
Calceolaria mocinoana Kuntze, Revis. gen. pl. 1: 41. 1891
Ionidium riparium A.Gray var. aestivum A.Gray, Pl. wright. 2: 16. 1852
Ionidium calceolarium Ging., in DC., Prodr. 1: 311. 1824
Ionidium parietariifolium DC. var. berterii DC., Prodr. 1: 308. 1824
Description:
Caulescent annual or biennial herb (in our region) or perennial subshrub, stem branched from lower nodes, erect from slender vertical rhizome with weakly branched roots, ≤ 6 dm tall; foliage and peduncles glabrate or puberulent to hirtellous; stipules very small, free, linear, entire; leaves many, spreading, lowest (sub)opposite, middle and upper alternate, blades broadly lanceolate (rarely linear-lanceolate), base cuneate, apex abruptly attenuate to attenuate, margins shallowly crenate or low-serrate to serrulate, ≤ 81 × 24 mm, petioles ≤ 1 cm; pedicels 3–20 mm; chasmogamous flower ≤ 12 mm; sepals linear-lanceolate to lance-triangular, attenuate; corolla whitish to violet with yellow throat; bottom petal about 3 × as long as others, blade greatly expanded, ovate to reniform; chasmogamous and cleistogamous capsules 3–5 mm long, ovoid with very short beak, green drying tan, unspotted; seeds ca. 1.7 × 1.5 mm, suborbicular and flattened, black.
Similar species:
Vegetatively superficially similar to Cubelium concolor but the lower leaves are usually opposite (rather than alternate throughout), and leaf blade margins are uniformly shallowly create to serrulate. The flowers differ in several features including a larger white to violet corolla with yellow throat (rather than greenish-white corolla) and a large spurred petal blade (rather than a short one only slightly longer than lateral and upper petals).
Ecology:
Seasonally moist soils, usually near water, along washes and in riparian areas and canyons.
Geographic distribution:
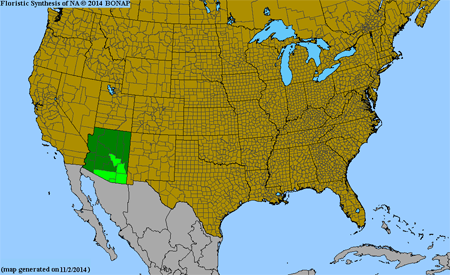
Southwestern U.S. (AZ) and Mexico, south to c. South America, introduced in tropical Asia; recently discovered by Keith Bradley and now documented as a well established introduction in citrus groves and along roadsides, and in two conservation areas, in Miami-Dade Co., FL, first vouchered by K. Bradley but reported incorrectly as P. linearifolia.
Conservation concern:
None.
Phenology:
Chasmogamous flower July–October, chasmogamous fruit July–November, cleistogamous fruit October.
Affinities:
Recent comprehensive phylogenetic investigations of the Violaceae family demonstrated that broadly circumscribed Hybanthus was extensively polyphyletic, and that the sole trait of a bottom petal which was saccate at base failed to delineate natural evolutionary groups (Feng 2005, Tokuoka 2008, Wahlert et al. 2014). The majority of New World hybanthoids formed a well supported clade and were subsequently segregated into the resurrected genus Pombalia on the basis of differences in floral and seed features and anatomical traits (Paula-Souza and Ballard 2014).
Hybrids:
None.
Comments:
Little and McKinney (2015) characterized this from southern Arizona, where it barely enters the U.S. from Mexico. This is the only (typically) broad-leaved member of Pombalia in our flora, and produces the largest flowers. The description was heavily supplemented with information by Little (2001).
Literature cited:
de Paula-Souza, J. and H. E. Ballard, Jr. 2014. Re-establishment of the name Pombalia, and new combinations from the polyphyletic Hybanthus (Violaceae). Phytotaxa 183(1): 1-15.
Feng, M. 2005. Floral morphogenesis and molecular systematics of the family Violaceae. PhD dissertation, Ohio University, Athens, OH. 279 pp.
Little, R. J. 2001. Violaceae Violet family. Journal of the Arizona-Nevada Academy of Science 33(1): 73-82.
Little, R. J., and L. E. McKinney. 2015. Violaceae. In Flora of North America: Cucurbitaceae to Droseraceae, 106. Oxford University Press, New York, NY.
Tokuoka, T. 2008. Molecular phylogenetic analysis of Violaceae (Malpighiales) based on plastid and nuclear DNA sequences. Journal of Plant Research 121(3): 253-260.
Wahlert, G. A., T. Marcussen, J. de Paula-Souza, M. Feng, and H. E. Ballard, Jr. 2014. A phylogeny of the Violaceae (Malpighiales) inferred from plastid DNA sequences: Implications for generic diversity and intrafamilial taxonomy. Systematic Botany 39: 239-252.

Habit by Max Licher, SERNEC portal

Chasmogamous flower by Gertrudes Yanes-Arvayo, SERNEC portal
Map by Biota of North America Program