Plasmodiophora

Neuhauser et al (2014) reported that based on 18s rDNA, Plasmodiophora diplantherae was in a clade within the Phagomyxida, distinct from P. brassicae in the Plasmodiophorida. Based on the molecular and morphological evidence, Hittorf et al (2020) reverted P. diplantherae to its original name Ostenfeldiella diplantherae Ferdinandsen and Winge within the Phagomyxida. We will keep the information about O. diplantherae here on the Plasmodiophora page, however, as an example of the limitations when using purely morphological data (viz., both optical and transmission electron microscopy) for classifications of taxa in the plasmodiphorids and phagomyxids. What is referred to as P. diplantherae in the following "Personal Comments" is now O. diplantherae. Similarly, in an extensive molecular and microscopic study, Kolátková et al (2023) reclassified what previously was known as P. bicaudata, along with a novel phytomyxid in root hairs of Zostera marina, into a new genus, Feldmanniella gen. nov., in a new, distinct clade in the Phagomyxida.
Personal Comments
After our success with the early ultrastructural descriptions of cruciform divisions in Sorosphaera and while Suzanne Harris was working on her ultrastructural karyotypes of Sorosphaera, I was on a sabbatical from Ohio University during 1978-79 in Michael Bennett's laboratory at the then Plant Breeding Institute (PBI), Trumpington, UK. Although my project was to work on meioisis in wheat, I was sidetracked a little because Paul Williams from the University of Wisconsin also was spending his sabbatical at PBI in another lab. Paul and I obviously had a lot to talk about concerning the plasmodiophorids, and we were fortunate to have in nearby Cambridge University, David Ingram. Paul, David and I decided to take a day trip to visit Stefan Buczacki at the then National Vegetable Research Station in Wellesborne, UK. That was when Stefan and I first met, and our meeting on that day led to a mini-sabbatical in his laboratory in Wellesborne during 1982 (more about Stefan is on the Tetramyxa page). I learned a lot that day about what Stefan, Paul, and David referred to as "Pb." After spending the day totally immerse in discussions about Pb, I was convinced that more basic ultrastructual research could be done on the plasmodiophorids in general.
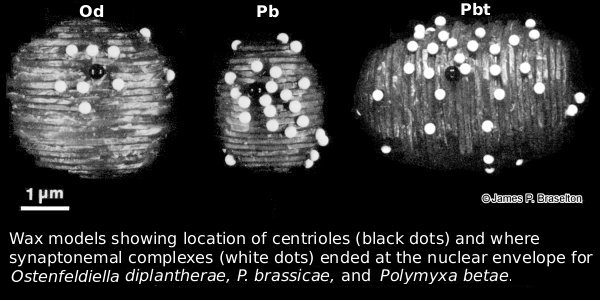
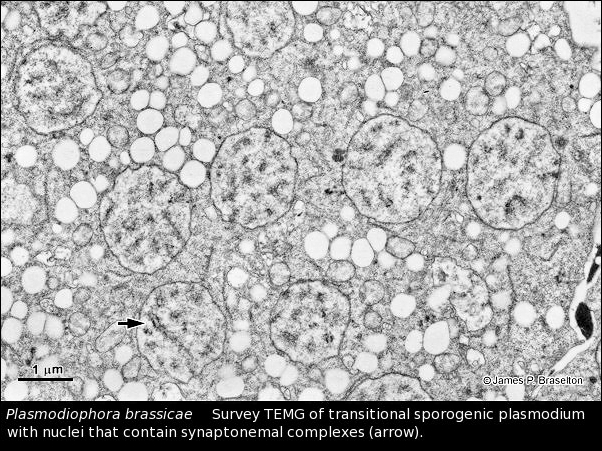
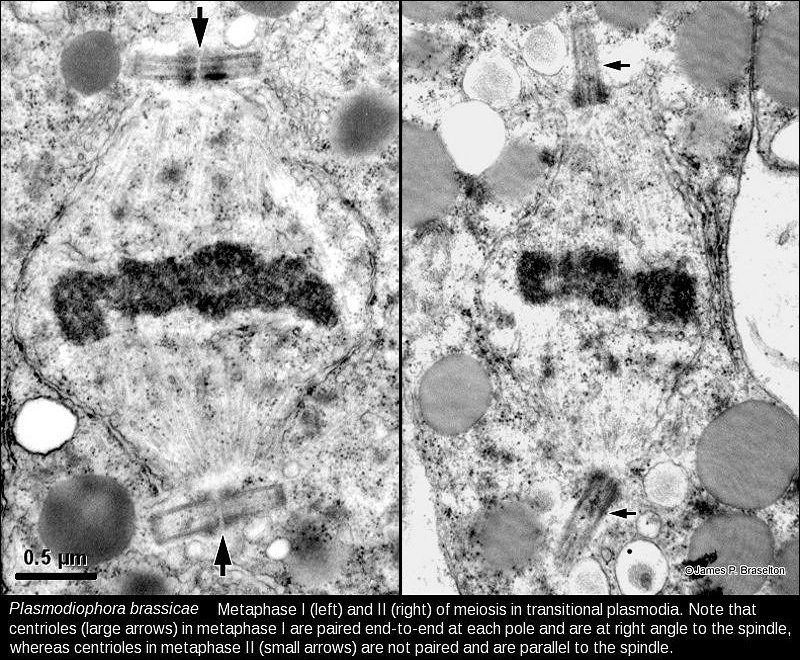
When I returned to Ohio University after the sabbatical and saw the success Suzanne Harris had with serial sectioning and karyotyping Sorosphaera, I set out to use the methods I learned on the sabbatical to karyotype as many plasmodiophorids as possible (see Meiosis & Karyotyping). Since Pb was the most economically significant member of the group and we knew meiosis with synaptonemal complexes occured in transitional sporogenic plasmodia (Garber & Aist 1979), it was the logical choice for the next karyotype of a plasmodiophorid.
Paul Williams had returned to Wisconsin from his sabbatical and was kind enough to send me inoculum of Pb. He also suggested that I take a trip up to the Ohio Agricultural Research and Development Center (OARDC) in Wooster to meet with Randy Rowe who was researching clubroot on Ohio radishes. Since I already had some friends in the electron microscopy center in plant pathology at the OARDC, it was an easy trip to arrange. Randy was very generous and provided additional inoculum in the form of clubbed radishes.
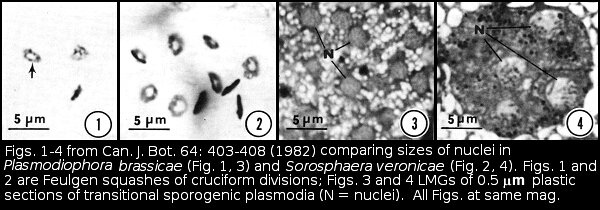
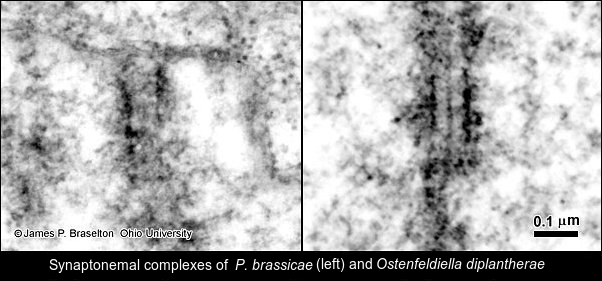
The synaptonemal complexes in Pb were difficult to follow in serial sections. It wasn't until I cut some very thick sections that I realized the trick to follow them was to use gold and almost purple sections, much thicker than the silver sections we generally used for transmission electron microscopy. I also was struck by how different in size Pb nuclei were from those of Sorosphaera.
{kind=link}
A few years later, after I observed that the karyotypes of the two species of Polymyxa were essentially identical, I thought it would be worthwhile to compare karyotypes of different species within another genus. Through a literature search I found that Frederick T. Short at the Jackson Estuarine Laboratory of the University of New Hampshire had researched shoal grass (Halodule wrightii), one of the hosts for what was known at the time as P. diplantherae. I wrote to Fred and asked if he had ever seen galls on plants in his study. He replied that he had. Through his assistance I was able to arrange for a collection trip to Harbor Branch Foundation, Inc., Fort Pierce, FL, in the spring of 1984. The date was easy to remember because the next day one of my sons, David, and I spent my 40th birthday at Disney World in Orlando. Another side note here: Fred and I have never met, but I remain indebted to him for introducing me to the folks at Harbor Branch Foundation.
Dennis Hanisak and Debbie Wescott at Harbor Branch Oceanographic Institute were generous with their time to show us the facilities. Debbie was our guide and took David and me out into the Indian River where I jumped into about 5 feet of water and pulled up Halodule that was loaded with galls. It wasn't until I was back in the boat that Debbie told me that there were a lot of manta rays in the area.
{kind=link}
{kind=link}
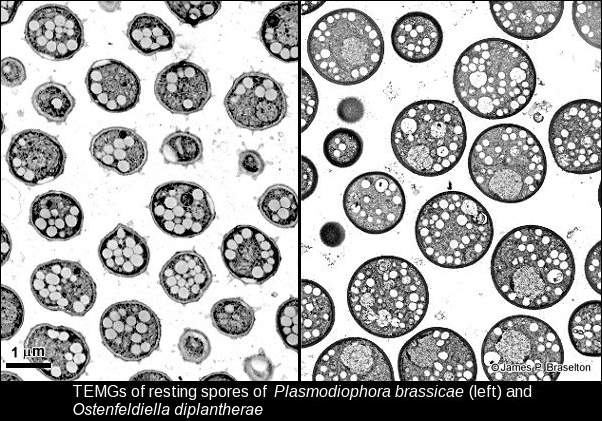
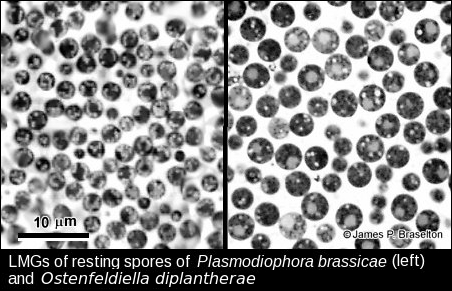
I was surprised at how different P. diplantherae was ultrastructurally from P. brassicae. Restings spores of P. diplantherae were larger than those of P. brassicae and had smooth walls without the ornamentation seen on resting spores of P. brassicae. The nuclei of of Pd were much larger than those of Pb, and the synaptonemal complexes much more clearly defined. Differences in length and number of the synaptonemal complexes were a total surprise. It is understandable that the molecular evidence has led to the renaming and reclassification of Plasmodiophora diplantherae to Ostenfeldiella diplantherae.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
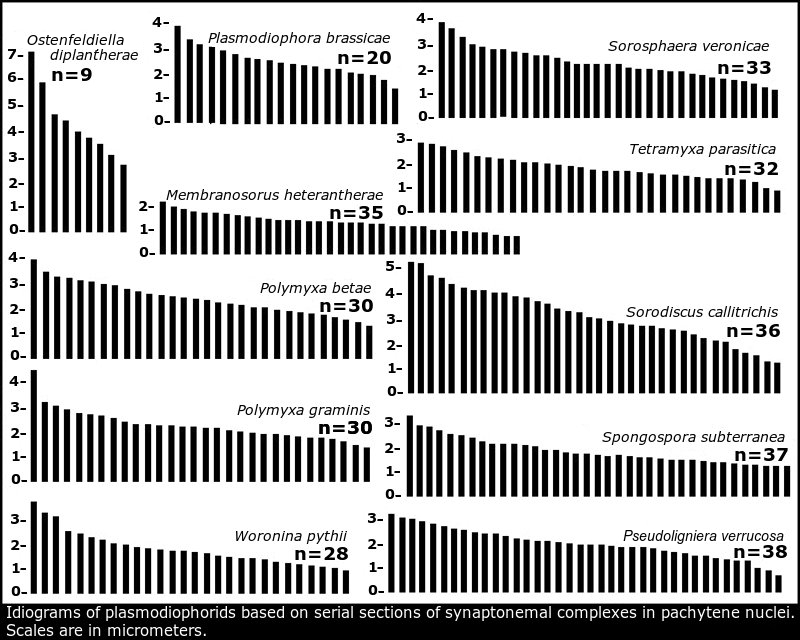
It was a pleasure to see that studies of the architecture of the P. brassicae nuclear and mitochondrial genomes (Stjelja et al 2019) and telomere-to-telomere genome assembly of P. brassicae (Javed et al 2024) came up with strikingly similar idiograms to the idiogram of the 20 chromosomes of P. brassicae based on serial sections of synaptonemal complexes by the present author (Braselton 1982) summarized in Idiograms based on synaptonemal complexes.
Images of Plasmodiophora and Ostenfeldiella
- Club root
- Molecular karyotype for P. brassicae
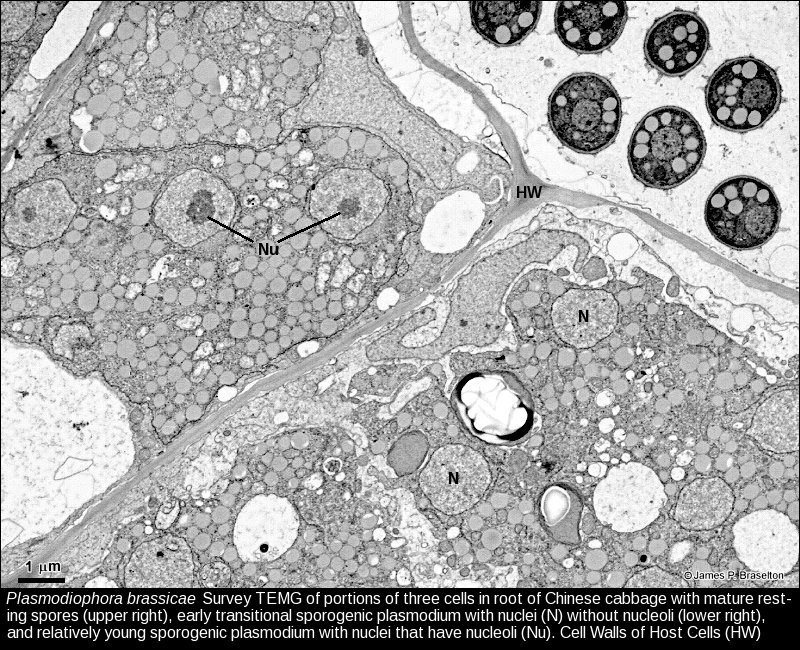
- Plasmodiophora sporogenic plasmodia, TEMG
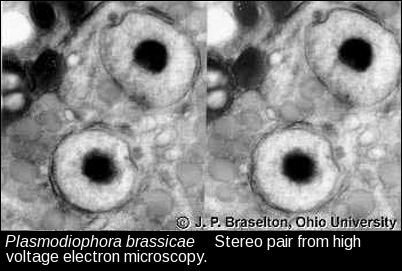
- Sporogenic interphase nuclei, HVTEM
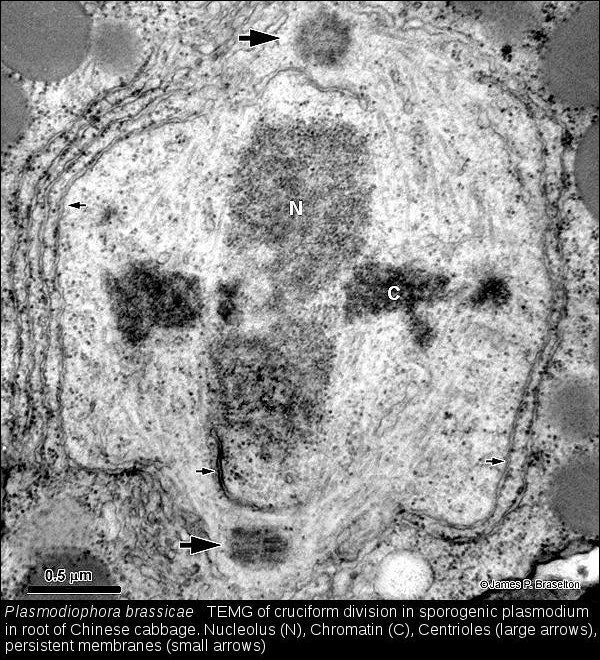
- Cruciform division of Plasmodiophora, TEMG
- Plasmodiophora sporogenic transitional plasmodium, TEMG
- Plasmodiophora metaphase I & II, TEMG
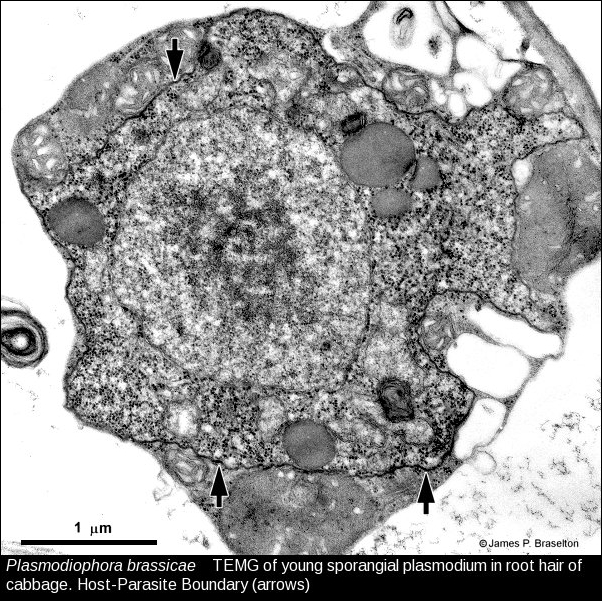
- Plasmodiophora young sporangial plasmodium, TEMG
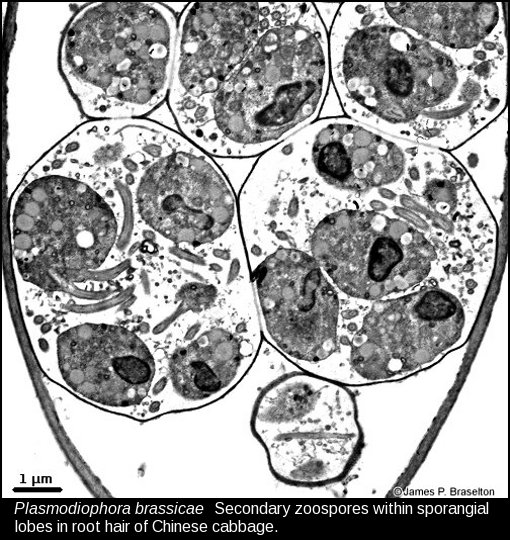
- Plasmodiophora sporangial lobes with zoospores, TEMG
- Plasmodiophora and Ostenfeldiella resting spores, LMG
- Plasmodiophora and Ostenfeldiella resting spores, TEMG
- Plasmodiophora and Ostenfeldiella synaptonemal complexes, TEMG
- Halodule infected with with Ostenfeldiella diplantherae
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Selected References for Plasmodiophora
There are a lot of references for Plasmodiophora. The following list is a sample of what is out there.
- Aist, J. R. & P. H. Williams. 1971. The cytology and kinetics of cabbage root hair penetration by Plasmodiophora brassicae Wor. Can. J. Bot. 49: 2023-2034.
- Ayers, G. W. 1944. Studies on the life history of the club root organism, Plasmodiophora brassicae. Can. J. Res. 22: 143-149.
- Braselton, J. P. 1982. Karyotypic analysis of Plasmodiophora brassicae based on serial thin sections of pachytene nuclei. Can. J. Bot. 60: 403-408.
- _____ & F. T. Short 1985. Karyotypic analysis of Plasmodiophora diplantherae. Mycologia 77: 940-945.
- Bulman, S., J. M. Candy, M. Fiers, R. Lister, A. J. Conner, & C.C. Eady. 2011. Genomics of biotrophic, plant-infecting plasmodiophorids using in vitro dual cultures. Protist 162: 449-461.
- Castlebury, J. A., J. V. Maddox, & D. A. Glawe. 1994. A technique for the extraction and purification of viable Plasmodiophora brassicae resting spores from host root tissue. Mycologia 86: 458-460.
- Dixon, G. R. 2009. The occurrence and economic impact of Plasmodiophora brassicae and clubroot disease. J. Plant Growth Regul. 28: 194-202.
- Dobson, R. L. & R. L. Gabrielson. 1983. Role of primary and secondary zoospores of Plasmodiophora brassicae in the development of clubroot in Chinese cabbage. Phytopathology 73: 559-561.
- Garber, R. C. & J. R. Aist. 1979. The ultrastructure of meiosis in Plasmodiophora brassicae (Plasmodiophorales). Can. J. Bot. 57: 2509-2518.
- Garber, R. C. & J. R. Aist. 1979. The ultrastructure of mitosis in Plasmodiophora brassicae (Plasmodiophorales). J. Cell Sci. 40: 89-110.
- Hartog, C. den. 1965. Some notes on the distribution of Plasmodiophora diplantherae, a parasitic fungus on species of Halodule. Persoonia 4: 15-18.
- Hittorf, M. et al. 2020. Revised taxonomy and expanded biodiversity of the Phytomyxea (Rhizaria, Endomyxa). J. Eukaryot. Microbiol. Doi: 10.1111/jeu.12817
- Ingram, D. S. & I. C. Tommerup. 1972. The life history of Plasmodiophora brassicae Woron. Proc. R. Soc. Lond. B. 180: 103-112.
- Javed, M. A. et al. 2024. Telomere-to-telomere genome assembly of the clubroot pathogen Plasmodiophora brassicae. Genome Biol. Evol. 16 Doi: 10.1093/gbe/evae122
- Kolátková, V. et al. 2023. Eelgrass (Zostera spp.) associated phytomyxids are host-specific congeneric parasites and predominant eukaryotes in the eelgrass rhizosphere on a global scale. Environ Microbiol. 25:1522–1537. Doi: 10.1111/1462-2920.16376
- Ludwig-Müller, J. & A. Schuller. 2008. What can we learn from clubroots: alterations in host roots and hormone homeostasis caused by Plasmodiophora brassicae. Eur. J. Plant Pathol. 121: 291-302.
- MacFarlane, I. 1952. Factors affecting the survival of Plasmodiophora brassicae Wor. in the soil and its assessment by a host test. Ann. Appl. Biol. 39: 239-256.
- MacFarlane, I. 1970. Germination of resting spores of Plasmodiophora brassicae. Trans. Br. mycol. Soc. 55: 97-112.
- Mithen, R. & R. Magrath. 1992. A contribution to the life history of Plasmodiophora brassicae: secondary plasmodia development in root galls of Arabidopsis thaliana. Mycol. Res. 96: 877-885.
- Niwa, R., A. Kawahara, H. Murakami, S. Tanakac, & T.Ezawa. 2011. Complete sructure [sic] of nuclear rDNA of the obligate plant parasite Plasmodiophora brassicae: Intraspecific polymorphisms in the exon and group I intron of the large subunit rDNA. Protist 162: 423-434.
- Schuller, A. & J. Ludwig-Müller. 2016. Histological methods to detect the clubroot pathogen Plasmodiophora brassicae during its complex life cycle. Plant Pathology 65: 1223-1237. Doi: 10.1111/ppa.12520
- Schwelm, A. et al. 2015 The Plasmodiophora brassicae genome reveals insights in its life cycle and ancestry of chitin synthases. Sci. Rep. 5: 11153. Doi: 10.1038/srep11153
- Scott, E. S. 1985. Production and characterization of single-spore isolates of Plasmodiophora brassicae. Plant Path. 34: 287-292.
- Stjelja, S., J. Fogelqvist, C. Tellgren-Roth, & C. Dixelius. 2019. The architecture of the Plasmodiophora brassicae nuclear and mitochondrial genomes. Scientific Reports 9:15753 Doi: 10.1038/s41598-019-52274-7
- Williams, P. H. & S. S. McNabola. 1967. Fine structure of Plasmodiophora brassicae in sporogenesis. Can. J. Bot. 45: 1665-1669.
- Williams, P. H. & S. S. McNabola. 1970. Fine structure of the host-parasite interface of Plasmodiophora brassicae in cabbage. Phytopathology 60: 1557-1561.
- Yukawa, Y. & S. Tanaka. 1979. Scanning electron microscope observations on resting sporangia of Plasmodiophora brassicae in clubroot tissues after alcohol cracking. Can. J. Bot. 57: 2528-2532.